 жқҗж–ҷдёҺж–№жі•
жқҗж–ҷдёҺж–№жі•
1.1 иҜ•йӘҢжқҗж–ҷ
иҸҢж ӘпјҡиҚҜзү©ж•Ҹж„ҹжҖ§иҜ•йӘҢиҙЁжҺ§иҸҢж ӘеӨ§иӮ жқҶиҸҢATCC 25922пјҢз”ұеұұдёңзңҒз•ңзҰҪз–«з—…йҳІжІ»дёҺз№ҒиӮІйҮҚзӮ№е®һйӘҢе®ӨдҝқеӯҳгҖӮ
и©ҰеүӮдёҺиҚҜе“Ғпјҡж°ЁиӢ„иҘҝжһ—гҖҒзҺҜдёҷжІҷжҳҹе’ҢеӨҡзІҳиҸҢзҙ пјҢиҙӯиҮӘдёӯеӣҪиҚҜе“Ғз”ҹзү©еҲ¶е“ҒжЈҖе®ҡжүҖпјҢеҲҶжһҗзәҜ;MHж¶ІдҪ“еҹ№е…»еҹәпјҲMHBпјүгҖҒMHеӣәдҪ“еҹ№е…»еҹәпјҲMHAпјүпјҢиҙӯиҮӘеҢ—дә¬йҷҶжЎҘжҠҖжңҜжңүйҷҗиҙЈд»»е…¬еҸёгҖӮ
1.2 иҜ•йӘҢж–№жі•
1.2.1 MICжөӢе®ҡ ж №жҚ®зҫҺеӣҪдёҙеәҠе’Ңе®һйӘҢе®Өж ҮеҮҶеҚҸдјҡпјҲClinical and Laboratory Standards InstituteпјҢCLSIпјүе»әз«Ӣзҡ„ж ҮеҮҶиҚҜж•Ҹж–№жі•пјҢйҮҮз”Ёеҫ®йҮҸиӮүжұӨзЁҖйҮҠжі•[9]пјҢжөӢе®ҡеҺҹе§ӢиҸҢж ӘгҖҒжҢҒз•ҷиҸҢеҸҠжҠ—иҚҜиҸҢж ӘеҜ№ж°ЁиӢ„иҘҝжһ—гҖҒзҺҜдёҷжІҷжҳҹгҖҒеӨҡзІҳиҸҢзҙ дёүз§ҚжҠ—иҸҢиҚҜзү©зҡ„жңҖе°ҸжҠ‘иҸҢжө“еәҰпјҲminimum inhibitory concentrationпјҢMICпјүгҖӮ
1.2.2 й«ҳиҚҜзү©жө“еәҰдёӢиҺ·еҫ—жҢҒз•ҷиҸҢж Ә з”Ёеҫ®йҮҸиӮүжұӨзЁҖйҮҠжі•жөӢе®ҡеӨ§иӮ жқҶиҸҢATCC 25922 MICеҗҺпјҢеңЁ96еӯ”жқҝдёҠпјҢе°Ҷж°ЁиӢ„иҘҝжһ—гҖҒзҺҜдёҷжІҷжҳҹе’ҢеӨҡзІҳиҸҢзҙ зҡ„иҚҜзү©жө“еәҰеҲҶеҲ«и®ҫзҪ®дёәеҗ„з§ҚиҚҜзү©MICзҡ„64еҖҚпјҲж°ЁиӢ„иҘҝжһ—256Ојg/mLпјҢзҺҜдёҷжІҷжҳҹ1Ојg/mLпјҢеӨҡзІҳиҸҢзҙ 32Ојg/mLпјүпјҢжҢүз…§иҚҜж•ҸиҜ•йӘҢж–№жі•зҡ„жҺҘз§ҚйҮҸжҺҘе…ҘзЁіе®ҡжңҹеӨ§иӮ жқҶиҸҢATCC 25922пјҢжҜҸз§ҚиҚҜеҒҡ18дёӘйҮҚеӨҚеӯ”пјҢ37в„ғйқҷзҪ®еҹ№е…»гҖӮеҲҶеҲ«дәҺ6 dеҶ…жҜҸеӨ©еҸ–3дёӘйҮҚеӨҚж ·пјҢе°ҶжҜҸеӯ”ж¶ІдҪ“е…ұ200ОјLе…ЁйғЁеҸ–еҮәпјҢ10 000 r/min зҰ»еҝғ5 minпјҢз”Ё200ОјLз”ҹзҗҶзӣҗж°ҙжҙ—ж¶Ө2ж¬ЎпјҢеҠ 50ОјLз”ҹзҗҶзӣҗж°ҙйҮҚжӮ¬еҗҺж¶ӮеёғMHAе№іжқҝпјҢеҹ№е…»24 пҪһ 48 hпјҢи®Ўз®—иҸҢиҗҪж•°пјҢжөӢе®ҡMICгҖӮ
1.2.3 еңЁйҖ’еўһзҡ„дәҡиҮҙжӯ»йҮҸиҚҜзү©жө“еәҰдёӢеҲҶжү№дј д»ЈиҺ·еҫ—жҠ—иҚҜиҸҢж Ә еҸ–еӨ§иӮ жқҶиҸҢATCC 25922жҺҘз§ҚдәҺиЈ…жңү50 mL MHBзҡ„дёү角瓶дёӯпјҲжҺҘз§ҚйҮҸдёә1%пјҢV/VпјүпјҢж°ЁиӢ„иҘҝжһ—гҖҒзҺҜдёҷжІҷжҳҹе’ҢеӨҡзІҳиҸҢзҙ зҡ„з»Ҳжө“еәҰеҲҶеҲ«дёәеҗ„иҮӘзҡ„1/2 MICпјҢ12 hеҗҺиҪ¬жҺҘеҲ°дёӢдёҖдёӘдёү角瓶дёӯпјҲиҚҜзү©жө“еәҰдёҚеҸҳпјүпјҢеҹ№е…»12 hеҗҺеҶҚиҪ¬жҺҘеҲ°иҚҜзү©жө“еәҰеҖҚеўһзҡ„дёү角瓶дёӯ;иҚҜзү©жө“еәҰз”ұ1/2 MICиҝһз»ӯеҖҚжҜ”йҖ’еўһиҮі64еҖҚMICпјҢжҜҸдёҖжө“еәҰдј д»Ј2ж¬ЎпјҢ第дәҢж¬Ўеҹ№е…»з»ҶиҸҢз”ҹй•ҝеҗҺеҲ’зәҝдәҺеҗ«зӣёеҗҢжө“еәҰиҚҜзү©зҡ„MHAе№іжқҝпјҢиҝҮеӨңеҹ№е…»пјҢжҢ‘еҸ–з”ҹй•ҝиүҜеҘҪзҡ„еҚ•иҸҢиҗҪ并жөӢе®ҡMICгҖӮ然еҗҺе°Ҷе…¶еңЁж— иҚҜMHBдёӯиҝһз»ӯдј д»ЈзӣҙиҮіMICзЁіе®ҡпјҢиҺ·еҫ—дёҖзі»еҲ—MICзЁіе®ҡзҡ„жҠ—иҚҜжҖ§еўһејәзҡ„иҸҢж ӘгҖӮ
1.2.4 жҢҒз•ҷиҸҢж ӘдёҺжҠ—иҚҜиҸҢж Әзҡ„з”ҹй•ҝжӣІзәҝжөӢе®ҡ е°ҶиҺ·еҫ—зҡ„еҺҹе§ӢиҸҢзҡ„жҢҒз•ҷиҸҢж Әе’ҢжҠ—иҚҜиҸҢж ӘеңЁеІӣжҙҘ2500 UV-VISе…үи°ұд»Әдёӯ37в„ғеҹ№е…»900 minпјҢжҜҸ15 minеңЁзәҝиҝһз»ӯжөӢйҮҸOD600пјҢжҜ”иҫғдёҚеҗҢиҸҢж Әзҡ„з”ҹй•ҝжӣІзәҝжҳҜеҗҰеӯҳеңЁе·®ејӮгҖӮ
1.2.5 жҢҒз•ҷиҸҢж Әе’ҢжҠ—иҚҜиҸҢж Әзҡ„MICиҢғеӣҙ еҲҶеҲ«йҖүеҸ–ж°ЁиӢ„иҘҝжһ—гҖҒзҺҜдёҷжІҷжҳҹе’ҢеӨҡзІҳиҸҢзҙ дёүз§ҚиҚҜзү©еҺҹе§ӢиҸҢзҡ„жҢҒз•ҷиҸҢж Әе’ҢжҠ—иҚҜиҸҢж ӘпјҢжҢ‘еҸ–еҚ•иҸҢиҗҪеҲҶеҲ«жҺҘз§ҚдәҺзӯүдҪ“з§Ҝзҡ„MHBдёӯпјҢд»ҘеҗҢж ·зҡ„жқЎд»¶жҢҜиҚЎеҹ№е…»16 пҪһ 18 hпјҢеҗ„еҸ–иҸҢж¶І1 mLзЁҖйҮҠиҮійҖӮеҪ“жө“еәҰпјҢж¶ӮеёғдәҺMHAе№іжқҝиҺ·еҫ—еҚ•иҸҢиҗҪпјҢеҗ„еҸ–100дёӘеҚ•иҸҢиҗҪиҝӣиЎҢMICжөӢе®ҡ[9]пјҢз»ҹи®ЎдёҚеҗҢиҸҢж Әеҗ„иҮӘзҡ„MICеҲҶеёғиҢғеӣҙгҖӮ
1.2.6 жҢҒз•ҷиҸҢж Әе’ҢжҠ—иҚҜиҸҢж Әз”ҹй•ҝдјҳеҠҝзҡ„жҜ”иҫғ ж №жҚ®1.2.5дёӯжөӢе®ҡзҡ„MICйў‘зҺҮеҲҶеёғз»“жһңпјҢйҖүеҸ–еҗ„иҮӘMICиҢғеӣҙжІЎжңүдәӨеҸүзҡ„жҢҒз•ҷиҸҢж Әе’ҢжҠ—иҚҜиҸҢж ӘпјҢзӯүжҜ”дҫӢжҺҘз§Қеҹ№е…»еҹәж··еҗҲеҹ№е…»пјҢд»Ҙеҗ„иҮӘMICдёәжЈҖжөӢжҢҮж Үз»ҹи®ЎдәҢиҖ…з»„жҲҗжҜ”дҫӢзҡ„еҸҳеҢ–гҖӮе…·дҪ“ж–№жі•пјҡжҢ‘еҸ–ж°ЁиӢ„иҘҝжһ—гҖҒзҺҜдёҷжІҷжҳҹе’ҢеӨҡзІҳиҸҢзҙ зҡ„жҢҒз•ҷиҸҢж Әе’ҢжҠ—иҚҜиҸҢж Әзҡ„еҚ•иҸҢиҗҪпјҢеҲҶеҲ«жҺҘз§ҚдәҺMHBдёӯпјҢ37в„ғгҖҒ200 r/minжҢҜиҚЎеҹ№е…»12 hпјҢз”ЁMHBзЁҖйҮҠпјҢдәҺзҙ«еӨ–-еҸҜи§Ғе…үеҲҶжһҗд»ӘпјҲGE Ultrospec 3100 proпјүжЈҖжөӢOD600пјҢи°ғиҠӮиҮі0.08 пҪһ 0.10иҢғеӣҙеҶ…пјҢе°ҪйҮҸи°ғж•ҙзЁҖйҮҠеҖҚж•°дҪҝе…¶жңҖз»ҲиҫҫеҲ°зӣёеҗҢзҡ„OD600еҖјпјҢзЎ®дҝқе°Ҹж•°зӮ№еҗҺдёӨдҪҚзӣёеҗҢгҖӮд»ҘжӯӨжө“еәҰеҗ„еҸ–500ОјLпјҢеҗҢж—¶жҺҘз§ҚдәҺеҗ«жңү100 mL MHBзҡ„дёү角瓶дёӯпјҢзӯүжҜ”дҫӢжҺҘз§ҚеҗҺж··еҗҲжҢҜиҚЎеҹ№е…»пјҢжҜҸеӨ©еҸ–1 mLиҪ¬жҺҘе…Ҙж–°зҡ„100 mL MHBдёӯпјҢиҝһз»ӯиҪ¬з§»дј д»Ј7 dз”ҡиҮіжӣҙй•ҝж—¶й—ҙгҖӮжҜҸж¬Ўеҹ№е…»еҗҺеҸ–ж··еҗҲиҸҢж¶ІзЁҖйҮҠиҮіеҗҲйҖӮжө“еәҰпјҢж¶ӮеёғиҺ·еҫ—еҚ•иҸҢиҗҪ100ж ӘпјҢжөӢе®ҡе…¶MICгҖӮд»Ҙ1.2.5дёӯжөӢе®ҡзҡ„MICйў‘зҺҮеҲҶеёғдёәжҢҮж ҮпјҢз»ҹи®ЎиҸҢж¶ІдёӯжҢҒз•ҷиҸҢж Әе’ҢжҠ—иҚҜиҸҢж ӘжқҘжәҗзҡ„еӨ§иӮ жқҶиҸҢзҡ„жҜ”дҫӢгҖӮ
2 з»“жһңдёҺеҲҶжһҗ
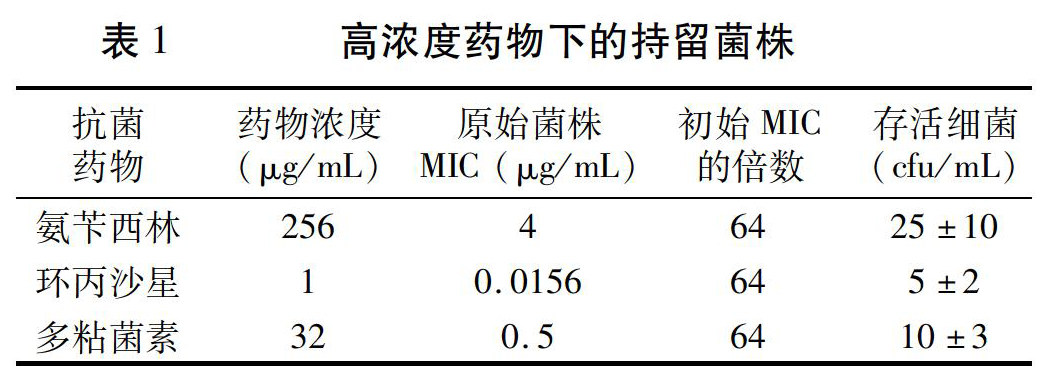
2.1 й«ҳиҚҜзү©жө“еәҰдёӢиҺ·еҫ—жҢҒз•ҷиҸҢж Ә
еңЁй«ҳиҚҜзү©жө“еәҰзҡ„еҹ№е…»жқЎд»¶пјҲж°ЁиӢ„иҘҝжһ—256Ојg/mLпјҢзҺҜдёҷжІҷжҳҹ1Ојg/mLпјҢеӨҡзІҳиҸҢзҙ 32Ојg/mLпјүдёӢпјҢжҢҒз»ӯдҪңз”Ё6 dпјҢдҫқ然иғҪеӨҹжЈҖжөӢеҲ°еӯҳжҙ»зҡ„з»ҶиҸҢгҖӮжҢүз…§жҺҘз§ҚйҮҸ104 cfu/mLж•°йҮҸзә§и®Ўз®—пјҢеӯҳжҙ»зҺҮеңЁ0.1%е·ҰеҸіпјҲиЎЁ1пјүгҖӮжӯӨиҜ•йӘҢз»“жһңиЎЁжҳҺпјҢеҚідҪҝеңЁжңүйҷҗзҡ„з©әй—ҙеҶ…пјҢеҸӘиҰҒеҲқе§ӢжҺҘз§ҚйҮҸиҫҫеҲ°дёҖе®ҡж•°йҮҸпјҢй«ҳдәҺиҮҙжӯ»жө“еәҰзҡ„жҠ—иҸҢиҚҜзү©е№¶дёҚиғҪе®Ңе…ЁжқҖжӯ»з»ҶиҸҢпјҢжҖ»дјҡжңүдёҖе°ҸйғЁеҲҶз»ҶиҸҢеӯҳжҙ»дёӢжқҘгҖӮ
еңЁиҚҜзү©ж°ЁиӢ„иҘҝжһ—жө“еәҰдёә256Ојg/mLжқЎд»¶дёӢпјҢе…ҲеҗҺе…ұ收йӣҶеӯҳжҙ»зҡ„з»ҶиҸҢ134ж ӘпјҢжөӢе®ҡе…¶MICпјҢ90%д»ҘдёҠзҡ„иҸҢж ӘдёҺеҺҹе§ӢиҸҢж ӘжҢҒе№іпјҢе°‘ж•°з•Ҙй«ҳдәҺеҺҹе§ӢиҸҢж ӘдёҖдёӘжҲ–дёӨдёӘжўҜеәҰпјҢеқҮиҝңиҝңдҪҺдәҺе…¶з”ҹеӯҳзҺҜеўғзҡ„иҚҜзү©жө“еәҰгҖӮзҺҜдёҷжІҷжҳҹе’ҢеӨҡзІҳиҸҢзҙ дҪңз”ЁдёӢзҡ„зҫӨдҪ“д№ҹеӯҳеңЁеҗҢж ·зҡ„зҺ°иұЎпјҲж•°жҚ®з•ҘпјүгҖӮиҝҷдәӣеңЁй«ҳжө“еәҰиҚҜзү©дҪңз”ЁдёӢеӯҳжҙ»зҡ„з»ҶиҸҢпјҢе…¶MICеҚҙдҝқжҢҒдёҚеҸҳпјҢдёәжҢҒз•ҷиҸҢгҖӮиҮҙжӯ»йҮҸзҡ„жҠ—иҸҢиҚҜзү©дёҚиғҪе°Ҷе…¶жқҖзҒӯпјҢиҜҙжҳҺжҢҒз•ҷиҸҢйҖҡиҝҮжҹҗз§ҚжңәеҲ¶иҖҗиҝҮдәҶй«ҳжө“еәҰзҡ„иҚҜзү©пјҢ并且дҝқжҢҒдәҶеҺҹжңүзҡ„иҚҜзү©ж•Ҹж„ҹзү№жҖ§гҖӮйҡҸжңәйҖүеҸ–е…¶дёӯдёҖж Әж°ЁиӢ„иҘҝжһ—зҡ„жҢҒз•ҷиҸҢпјҢеңЁж°ЁиӢ„иҘҝжһ—жө“еәҰдёә256Ојg/mLзҡ„зҺҜеўғдёӯеҫ—д»Ҙеӯҳжҙ»пјҢжөӢе®ҡе…¶еҲҶиЈӮз№Ғж®–зҡ„еҗҺд»Је®һйҷ…MICдёә4Ојg/mLпјҢз»Ҹдј д»ЈеҗҺMICдҝқжҢҒдёҚеҸҳпјҢе‘ҪеҗҚдёәиҸҢж ӘP-AпјҢжҳҜеҺҹе§ӢиҸҢж ӘжҢҒз•ҷиҸҢж Әз№Ғж®–еҗҺд»Јзҡ„зәҜеҹ№е…»гҖӮ
з”ЁеҗҢж ·зҡ„ж–№жі•иҺ·еҫ—зҺҜдёҷжІҷжҳҹе’ҢеӨҡзІҳиҸҢзҙ дҪңз”ЁдёӢзҡ„жҢҒз•ҷиҸҢж ӘпјҢеҲҶеҲ«иҖҗиҝҮзҡ„иҚҜзү©жө“еәҰдёә1Ојg/mLе’Ң32Ојg/mLпјҢдҝқжҢҒеҺҹе§ӢиҸҢж Әзҡ„MICеҲҶеҲ«дёә0.0156Ојg/mLе’Ң0.5Ојg/mLдёҚеҸҳгҖӮ
2.2 еңЁйҖ’еўһзҡ„дәҡиҮҙжӯ»йҮҸиҚҜзү©жө“еәҰдёӢдј д»ЈиҺ·еҫ—жҠ—иҚҜиҸҢж Ә
еңЁиҚҜзү©йҖүжӢ©еҺӢеҠӣдёҚж–ӯжҸҗй«ҳзҡ„иҜ•йӘҢжқЎд»¶дёӢиҺ·еҫ—дёҖзі»еҲ—жҠ—иҚҜжҖ§еўһејәзҡ„иҸҢж ӘпјҢиғҪеӨҹеңЁзӣёеә”иҚҜзү©жө“еәҰдёӢ继з»ӯз”ҹй•ҝз№Ғж®–гҖӮиҝҷдәӣиҸҢж Әзҡ„MICеҖјиҝңиҝңй«ҳдәҺеҺҹе§ӢиҸҢж ӘMICгҖӮз”ұдәҺеӯҳеңЁеӨҡз§ҚеҪўжҲҗжңәеҲ¶пјҢеңЁиҚҜзү©еҺ»йҷӨеҗҺжңүдәӣжҠ—иҚҜжңәеҲ¶е…ій—ӯпјҢиҖҢжңүдәӣжҠ—иҚҜжңәеҲ¶жҳҜзЁіе®ҡзҡ„пјҢеӣ жӯӨпјҢеңЁж— иҚҜеҹ№е…»еҹәдёӯиҝһз»ӯдј д»ЈеҗҺпјҢжҠ—иҚҜиҸҢж Әзҡ„MICеҖјжңүдёҚеҗҢзЁӢеәҰзҡ„еӣһиҗҪпјҢдҪҶйғҪйҡҫд»ҘеӣһеӨҚеҲ°еҺҹе§ӢиҸҢж Әзҡ„ж•Ҹж„ҹжҖ§гҖӮ
з»ҸйҖ’еўһзҡ„дәҡиҮҙжӯ»йҮҸж°ЁиӢ„иҘҝжһ—иҝһз»ӯеҹ№е…»еҗҺпјҢйҖүеҸ–дёҖж ӘMICзЁіе®ҡзҡ„иҖҗиҝҮж°ЁиӢ„иҘҝжһ—256Ојg/mLзҡ„жҠ—иҚҜиҸҢж ӘпјҢдј д»ЈеҗҺMICзЁіе®ҡеңЁ128Ојg/mLпјҢжҳҜеҺҹе§ӢиҸҢж ӘMICзҡ„32еҖҚпјҢе‘ҪеҗҚдёәR-AгҖӮ
з”ЁеҗҢж ·зҡ„ж–№жі•иҺ·еҫ—зҺҜдёҷжІҷжҳҹе’ҢеӨҡзІҳиҸҢзҙ зҡ„жҠ—иҚҜиҸҢж ӘпјҢйҖүеҸ–иҖҗиҝҮ1Ојg/mLзҺҜдёҷжІҷжҳҹе’Ң32Ојg/mLеӨҡзІҳиҸҢзҙ зҡ„жҠ—иҚҜиҸҢж ӘпјҢзЁіе®ҡзҡ„MICеҲҶеҲ«дёә0.5Ојg/mLе’Ң8Ојg/mLгҖӮ
2.3 жҢҒз•ҷиҸҢж Әе’ҢжҠ—иҚҜиҸҢж Әзҡ„MICиҢғеӣҙ
MICеҖјиЎЁзӨәиҸҢж ӘеҜ№иҚҜзү©зҡ„ж•Ҹж„ҹзЁӢеәҰпјҢжҳҜдёҖдёӘзҫӨдҪ“жҰӮеҝөгҖӮз”ұдәҺз»ҶиҸҢзҫӨдҪ“е…·жңүејӮиҙЁжҖ§пјҢзәҜеҹ№е…»зҡ„з»ҶиҸҢзҫӨдҪ“дёӯе°ҸдәҡзҫӨзҡ„MICд№ҹеҸҜиғҪжңүжүҖе·®ејӮгҖӮеӣ жӯӨпјҢMIC并йқһдёҖдёӘеҮҶзЎ®зҡ„ж•°еҖјпјҢиҖҢжҳҜдёҖдёӘиҢғеӣҙгҖӮдёәдәҶеҗҺз»ӯжҜ”иҫғз”ҹй•ҝдјҳеҠҝпјҢе…ҲжЈҖжөӢеҗ„зұ»иҸҢж Әзҡ„MICиҢғеӣҙпјҢеҲҶеҲ«еҸ–100дёӘеҚ•иҸҢиҗҪжөӢе®ҡMICеҲҶеёғжғ…еҶөгҖӮж°ЁиӢ„иҘҝжһ—иҚҜзү©еӨ„зҗҶзҡ„иҸҢж ӘMICз»ҹи®Ўз»“жһңеҰӮиЎЁ2пјҢP-Aзҡ„MICеҲҶеёғеі°еҖјдёә4Ојg/mLпјҢMICдёҺеҺҹе§ӢиҸҢж ӘжҢҒе№і;R-Aзҡ„MICеҲҶеёғеі°еҖјдёә128Ојg/mLпјҢжҳҜеҺҹе§ӢиҸҢж ӘMICзҡ„32еҖҚгҖӮз»ҸиҝҮдј д»Јеҹ№е…»иҜҒжҳҺпјҢдёӨдёӘиҸҢж Әзҡ„MICиҢғеӣҙжҳҜзЁіе®ҡзҡ„иЎЁеһӢжҢҮж ҮпјҢеҸҜд»ҘдҪңдёәеҲӨе®ҡиҸҢж ӘжҖ§иҙЁзҡ„жҢҮж ҮгҖӮеҜ№зҺҜдёҷжІҷжҳҹе’ҢеӨҡзІҳиҸҢзҙ еӨ„зҗҶиҸҢж Әзҡ„зЁіе®ҡMICиҢғеӣҙд№ҹжҢүеҗҢж ·зҡ„ж–№жі•иҝӣиЎҢз»ҹи®ЎпјҲж•°жҚ®з•ҘпјүгҖӮ
2.4 жҢҒз•ҷиҸҢж Әе’ҢжҠ—иҚҜиҸҢж Әз”ҹй•ҝдјҳеҠҝзҡ„жҜ”иҫғ
з»ҸжЈҖжөӢпјҢдёҚеҗҢиҚҜзү©дҪңз”ЁдёӢзӯӣйҖүзҡ„дёӨзұ»иҸҢж Әзҡ„з”ҹй•ҝжӣІзәҝе№¶ж— жҳҺжҳҫе·®ејӮпјҲз”ҹй•ҝжӣІзәҝеӣҫз•ҘпјүпјҢеӣ жӯӨеҸҜйҮҮеҸ–ж··еҗҲеҹ№е…»зҡ„ж–№ејҸпјҢд»Ҙеҗ„иҮӘMICдёәиЎЎйҮҸжҢҮж ҮжҜ”иҫғе…¶з”ҹй•ҝдјҳеҠҝгҖӮ
ж°ЁиӢ„иҘҝжһ—еӨ„зҗҶзҡ„P-Aе’ҢR-AиҸҢж ӘMICиҢғеӣҙж— зӣёдә’дәӨеҸүпјҢе°ҶдәҢиҖ…зӯүжҜ”дҫӢж··еҗҲжҺҘз§Қеҹ№е…»пјҢж №жҚ®зЁіе®ҡзҡ„MICиҢғеӣҙз»ҹи®Ўе…¶жҜ”дҫӢеҸҳеҢ–гҖӮеҰӮеӣҫ1жүҖзӨәпјҢж··еҗҲеҹ№е…»еҗҺпјҢжҢҒз•ҷиҸҢж ӘP-AжүҖеҚ жҜ”дҫӢйҖҗжёҗеҚҮй«ҳпјҢеҲ°з¬¬4 dеўһиҮі70%д»ҘдёҠпјҢиҮіз¬¬7 dжҠ—иҚҜиҸҢж ӘR-AжҜ”дҫӢдҪҺдәҺ5%пјҢиҸҢж ӘP-Aж•°йҮҸеҚ з»қеҜ№дјҳеҠҝгҖӮжӯӨз»“жһңиЎЁжҳҺпјҢеңЁжІЎжңүиҚҜзү©зҡ„еҹ№е…»жқЎд»¶дёӢпјҢP-Aе…·жңүжӣҙејәзҡ„з”ҹй•ҝдјҳеҠҝгҖӮйҡҸжңәжҢ‘йҖүж°ЁиӢ„иҘҝжһ—дҪңз”ЁдёӢеӯҳжҙ»зҡ„е…¶е®ғжҢҒз•ҷиҸҢж Әе’ҢжҠ—иҚҜиҸҢж ӘпјҢз”ЁеҗҢж ·зҡ„ж–№жі•еҜ№жҜ”е…¶з”ҹй•ҝдјҳеҠҝпјҢд№ҹжңүзұ»дјјзҡ„з»“жһңпјҲж•°жҚ®з•ҘпјүгҖӮеҸҰеӨ–дёӨз§ҚиҚҜзү©зҺҜдёҷжІҷжҳҹе’ҢеӨҡзІҳиҸҢзҙ зҡ„жҢҒз•ҷиҸҢж Әе’ҢжҠ—иҚҜиҸҢж ӘпјҢз”ҹй•ҝдјҳеҠҝзҡ„жҜ”иҫғд№ҹеӯҳеңЁзұ»дјјз»“жһңпјҲж•°жҚ®з•ҘпјүпјҢжңҖз»ҲйғҪиғҪдҪ“зҺ°еҮәжҢҒз•ҷиҸҢж Әе…·жңүжӣҙејәзҡ„з”ҹй•ҝдјҳеҠҝгҖӮжң¬з ”究дёӯе…ұжЈҖжөӢ8з»„иҸҢж Әзҡ„еҜ№жҜ”пјҢжңӘжЈҖжөӢеҲ°дҫӢеӨ–жғ…еҶөгҖӮ
3 и®Ёи®әдёҺз»“и®ә
3.1 з»ҶиҸҢжҠ—иҚҜжҖ§зҡ„ж„Ҹд№ү
жҠ—иҸҢиҚҜзү©зҡ„е№ҝжіӣеә”з”ЁжҳҜзӯӣйҖүдёҺеҜҢйӣҶжҠ—иҚҜиҸҢж Әзҡ„дё»иҰҒжҺЁеҠЁеҠӣпјҢеҚҙ并йқһжҠ—иҚҜжҖ§дә§з”ҹзҡ„ж №жәҗ[10]гҖӮжҠ—иҚҜеҹәеӣ е№ҝжіӣиҖҢжҷ®йҒҚең°еӯҳеңЁдәҺзҺҜеўғдёӯвҖ”з—…еҺҹиҸҢгҖҒе…ұз”ҹиҸҢеҸҠеӨҡз§ҚзҺҜеўғеҫ®з”ҹзү©[11]гҖӮи¶ҠжқҘи¶ҠеӨҡзҡ„з ”з©¶иЎЁжҳҺпјҢжҠ—иҚҜжҖ§жҳҜз»ҶиҸҢзҡ„дёҖдёӘиҮӘ然еұһжҖ§[12]пјҢеҪ»еә•ж¶ҲйҷӨжҠ—иҚҜжҖ§жҳҜдёҚеҸҜиғҪзҡ„гҖӮз”ұдәҺз»ҶиҸҢйҒ—дј зү©иҙЁзҡ„иҮӘеҸ‘зӘҒеҸҳгҖҒиҚҜзү©еҺӢеҠӣзҡ„зӯӣйҖүе’ҢиҜұеҜјгҖҒеӨҚжқӮеӨҡж ·зҡ„жҠ—иҚҜжҖ§жңәеҲ¶гҖҒдёҚеҸҜйҒҝе…Қең°дёҺе‘ЁеӣҙзҺҜеўғеҫ®з”ҹзү©жҺҘи§Ұзӯүдј—еӨҡеӣ зҙ пјҢеҚідҪҝеңЁжңүйҷҗзҡ„з©әй—ҙеҶ…е’Ңзү№е®ҡжўқ件дёӢд№ҹйҡҫд»Ҙе®Ңе…Ёж¶ҲйҷӨз»ҶиҸҢжҠ—иҚҜжҖ§гҖӮд»ҺеҸҰдёҖдёӘж–№йқўзңӢпјҢжҠ—иҚҜжҖ§еҜ№дәҺз»ҶиҸҢжң¬иә«иҖҢиЁҖеҲҷж„Ҹе‘ізқҖзү©з§ҚиғҪеӨҹе…ӢжңҚзҺҜеўғжҠ—иҸҢиҚҜзү©еҺӢеҠӣеҫ—д»Ҙ延з»ӯе’Ңеӯҳжҙ»пјҢеҜ№зҺҜеўғеҫ®з”ҹзү©зҫӨиҗҪз»“жһ„зҡ„еӨҡж ·жҖ§е’ҢзЁіе®ҡжҖ§иө·еҲ°дәҶйҮҚиҰҒдҪңз”ЁпјҢеҜ№еҠЁзү©пјҲеҢ…жӢ¬дәәпјүе…Қз–«зі»з»ҹзҡ„еҪўжҲҗгҖҒеҒҘеә·д№ғиҮіж•ҙдёӘз”ҹжҖҒзҺҜеўғйғҪе…·жңүйҮҚеӨ§еҪұе“Қ[13пјҢ14]гҖӮ
зҙ°иҸҢзҡ„жҠ—иҚҜжҖ§дёәиҸҢж ӘжҸҗдҫӣдәҶйўқеӨ–зҡ„з”ҹзү©еӯҰзү№жҖ§пјҢеё®еҠ©жҸҗй«ҳеҫ®з”ҹзү©еҜ№зү№е®ҡзҺҜеўғзҡ„йҖӮеә”жҖ§пјҢеҗҢж—¶еӨҡж•°жғ…еҶөдёӢдјҡд»ҳеҮәдёҖе®ҡзҡ„жҠ—жҖ§д»ЈеҒҝпјҢеҢ…жӢ¬з”ҹй•ҝз№Ғж®–иғҪеҠӣйҷҚдҪҺгҖӮеӣ жӯӨпјҢзҗҶи®әдёҠеҺҹе§ӢиҸҢж ӘжҜ”жҠ—иҚҜиҸҢж Әе…·жңүжӣҙејәзҡ„з”ҹй•ҝдјҳеҠҝпјҢдҪҶеңЁжҠ—иҸҢиҚҜзү©еҺӢеҠӣдёӢе…·жңүжӣҙејәз”ҹй•ҝдјҳеҠҝзҡ„еҺҹе§Ӣж•Ҹж„ҹиҸҢиў«жқҖжӯ»пјҢиҖҢжҠ—иҚҜиҸҢж Әзҡ„ж•Ҹж„ҹжҖ§еӣһеӨҚеҲҷжҳҜеӨҚжқӮжј«й•ҝзҡ„иҝҮзЁӢ[15]гҖӮ
3.2 жҢҒз•ҷиҸҢзҡ„еӯҳеңЁеҸҠеңЁжҠ—иҚҜжҖ§еӣһеӨҚдёӯзҡ„жҪңеҠӣ
еңЁжһ—жһ—жҖ»жҖ»зҡ„жҠ—иҚҜиЎЁеһӢе’ҢжҠ—иҚҜжңәеҲ¶дёӯпјҢжҢҒз•ҷиҸҢд»Ҙе…¶зӢ¬зү№зҡ„жңәеҲ¶еј•иө·дәә们зҡ„жіЁж„ҸгҖӮжҢҒз•ҷиҸҢйҖҡиҝҮвҖңдј‘зң -жҒўеӨҚз”ҹй•ҝ-еўһж®–вҖқзҡ„ж–№ејҸз»ҙжҢҒиҮӘиә«зҡ„з”ҹеӯҳе’ҢиҸҢдҪ“з»“жһ„зЁіе®ҡпјҢе·§еҰҷиәІиҝҮеӨ–з•ҢжҒ¶еҠЈзҺҜеўғзҡ„еҪұе“ҚиҖҢеӯҳз•ҷдёӢжқҘпјҢдҪҶдҫқ然дҝқжҢҒзқҖеҺҹе§ӢиҸҢж Әзҡ„зү№жҖ§гҖӮжҜ”иҫғжҢҒз•ҷиҸҢж ӘдёҺжҠ—иҚҜиҸҢж Әзҡ„з”ҹй•ҝдјҳеҠҝпјҢеҸҜеј•еҸ‘еҜ№жҠ—иҚҜжҖ§еӣһеӨҚй—®йўҳзҡ„йҮҚж–°жҖқиҖғгҖӮжң¬ж–Үз ”з©¶з»“жһңиЎЁжҳҺеҪ“еҺ»йҷӨжҠ—иҸҢиҚҜзү©еҺӢеҠӣеҗҺпјҢжҠ—иҸҢиҚҜзү©еҺӢеҠӣдёӢеӯҳжҙ»зҡ„жҢҒз•ҷиҸҢж ӘжҜ”жҠ—иҚҜиҸҢж Әе…·жңүжӣҙејәзҡ„з”ҹй•ҝдјҳеҠҝпјҢиҝҷдёҖз»“и®әе°ҡжңӘи§Ғе…¶е®ғзұ»дјјж–ҮзҢ®гҖӮиҷҪ然жҢҒз•ҷиҸҢзҡ„дҪңз”Ёе°ҡжңӘз»ҸиҝҮе…ЁйқўйӘҢиҜҒпјҢдҪҶйқўеҜ№з”ҹжҖҒдёӯдёҘеі»зҡ„жҠ—иҚҜжҖ§й—®йўҳпјҢжҢҒз•ҷиҸҢзҡ„еӯҳеңЁе’Ңзү№жҖ§иҝҳжҳҜжҸҗдҫӣдәҶдёҖзәҝеёҢжңӣгҖӮеңЁзҺҜеўғдёӯжҠ—иҸҢиҚҜзү©еҺӢеҠӣдёҚж–ӯеўһеӨ§зҡ„иҝҮзЁӢдёӯпјҢеӯҳз•ҷдәҶдёҖжү№жү№вҖңжІүзқЎвҖқзҡ„вҖңз§ҚеӯҗвҖқвҖ”вҖ”жҢҒз•ҷиҸҢгҖӮеҪ“иҚҜзү©еҺӢеҠӣйҖҗжёҗйҷҚдҪҺпјҢдәә们еҜ№дәҺжҠ—иҚҜжҖ§ж°ҙе№ійҷҚдҪҺзҡ„жңҹеҫ…дёҚд»…еҒңз•ҷеңЁжҠ—иҚҜиҸҢж Әзҡ„ж•Ҹж„ҹжҖ§жҒўеӨҚ[10]пјҢд№ҹеңЁдәҺйў„з•ҷзҡ„жҢҒз•ҷиҸҢвҖңиӢҸйҶ’вҖқ并жҒўеӨҚжҙ»еҠӣпјҢд»ҘжӣҙдёәеҺҹе§Ӣзҡ„иҸҢж Әзү№жҖ§дёҺзҺ°еӯҳзҡ„жҠ—иҚҜжҖ§з»ҶиҸҢе…ұеҗҢз«һдәүпјҢиҖҢеӨҡж•°жҠ—иҚҜжҖ§з»ҶиҸҢеҸҜиғҪз”ұдәҺе…¶жҠ—жҖ§д»ЈеҒҝиҖҢеӨ„дәҺеҠЈеҠҝгҖӮеҸӘжңүзҺҜеўғдёӯиҸҢзҫӨзҡ„жҠ—иҚҜжҖ§ж°ҙе№іж•ҙдҪ“еӣһиҗҪжүҚиғҪж №жң¬и§ЈеҶіжҠ—иҚҜжҖ§й—®йўҳпјҢ并йқһеҜ№жҠ—иҚҜиҸҢж–©е°ҪжқҖз»қпјҢд№ҹйқһйҳ»ж–ӯжүҖжңүжҠ—иҚҜжҖ§жңәеҲ¶гҖӮжң¬з ”究结и®әиЎЁжҳҺпјҢжҢҒз•ҷиҸҢзҡ„еӯҳеңЁе’ҢеҪўжҲҗжңәеҲ¶дҝқз•ҷдәҶеҺҹе§ӢиҸҢж Әзҡ„з”ҹй•ҝдјҳеҠҝпјҢйҡҸзқҖжҠ—иҸҢиҚҜзү©дҪҝз”Ёж°ҙе№ізҡ„йҷҚдҪҺпјҢеҸҜиғҪеңЁжҠ—иҚҜжҖ§зҡ„йҖҶиҪ¬дёӯиө·еҲ°дёҖе®ҡдҪңз”ЁгҖӮ
еҸӮ иҖғ ж–Ү зҢ®пјҡ
[1]Laxminarayan RпјҢ Duse AпјҢ Wattal CпјҢ et al. Antibiotic resistance - the need for global solutions[J]. The Lancet Infectious DiseasesпјҢ 2013пјҢ 13пјҲ12пјүпјҡ 1057-1098.
[2]Wax R GпјҢ Lewis KпјҢ Salyers A AпјҢ et al. Bacterial resistance to antimicrobials[M]. Second Edition. New Yorkпјҡ CRC PressпјҢ 2007.
[3]Kuehn B M. Antibiotic-resistant вҖңsuperbugsвҖқ may be transmitted from animals to humans[J]. The Journal of American Medical AssociationпјҢ 2007пјҢ 298пјҲ18пјүпјҡ 2125-2126.
[4]Bigger J W. Treatment of staphylococcal infections with penicillin by intermittent sterilization[J]. The LancetпјҢ 1944пјҢ 244пјҲ6320пјүпјҡ 497-500.
[5]Balaban N QпјҢ Merrin JпјҢ Chait RпјҢ et al. Bacterial persistence as a phenotypic switch[J]. ScienceпјҢ 2004пјҢ 305пјҲ5690пјүпјҡ1622-1625.
[6]Moyed H SпјҢ Bertrand K P. hipAпјҢ a newly recognized gene of Escherichia coli K-12 that affects frequency of persistence after inhibition of murein synthesis[J]. Jorunal of BacteriologyпјҢ 1983пјҢ 155пјҲ2пјүпјҡ 768-775.
[7]Gerdes KпјҢMaisonneuve E. Bacterial persistence and toxin-antitoxin loci[J]. The Annual Review of MicrobiologyпјҢ2012пјҢ66пјҡ103-123.
[8]Pu YпјҢ Zhao ZпјҢ Li YпјҢ et al. Enhanced efflux activity facilitates drug tolerance in dormant bacterial cells[J]. Molecular CellпјҢ 2016пјҢ 62пјҲ2пјүпјҡ 284-294.
[9]CLSI. Performance standards for antimicrobial disk susceptibility tests. 27th edition. CLSI document M100[S]. Wayneпјҡ Clinical and Laboratory Standards InstituteпјҢ 2017.
[10]Hughes D. Selection and evolution of resistance to antimicrobial drugs[J]. IUBMB LifeпјҢ 2014пјҢ 66пјҲ8пјүпјҡ 521-529.
[11]Allen H KпјҢ Donato JпјҢ Wang H HпјҢ et al. Call of the wildпјҡ antibiotic resistance genes in natural environments[J]. Nature Reviews MicrobiologyпјҢ 2010пјҢ 8пјҲ4пјүпјҡ 251-259.
[12]Martinez J L. The role of natural environments in the evolution of resistance traits in pathogenic bacteria[J]. Proceedings of the Royal Society Bпјҡ Biological SciencesпјҢ 2009пјҢ 276пјҲ1667пјүпјҡ 2521-2530.
[13]Sommer M O AпјҢ Dantas GпјҢ Church G M. Functional characterization of the antibiotic resistance reservoir in the human microfiora[J]. ScienceпјҢ 2009пјҢ 325пјҲ5944пјүпјҡ 1128-1131.
[14]Whitman W BпјҢ Coleman D CпјҢ Wiebe W J. Prokaryotesпјҡ the unseen majority[J]. Proceedings of the National Academy of Sciences of the United States of AmericaпјҢ 1998пјҢ 95пјҲ12пјүпјҡ 6578-6583.
[15]Andersson D IпјҢ Hughes D. Antibiotic resistance and its costпјҡ is it possible to reverse resistanceпјҹ[J]. Nature Reviews MicrobiologyпјҢ 2010пјҢ 8пјҲ4пјүпјҡ 260-271.
жҺЁиҚҗи®ҝй—®: